
“蜜蜂梦见了花朵,花朵梦见了蜜蜂。”弗朗西斯·赫斯黎(Francis Huxley)
“蜜蜂和花朵是在一起的,如果你拿走其中一个,二者都会消失。”弗朗西斯科·瓦雷拉(Francisco Varela)
在生物科学和计算机科学的交叉点上,在神经科学领域中盛行了五十多年的大脑与计算机隐喻导致孤立的个体思考,而没有考虑到个体的社会互动。现在,被称为超扫描的神经成像技术使我们能同时记录多个参与者的行为和大脑活动,从而研究无意识和对等条件下的社会互动。透过将两个个体视为单一系统,观察行为以及大脑活动,以整体的方式来理解人类的互动。这些方法论和理论的发展证明了与他人的互动和被动消极的社会知觉有著根本的不同。这些结果促使我们在神经科学工作中更广泛地思考互动的尺度,以及我们的社会互动和生物学基础之间的互补性。
从控制论到自组织,用哪种典范来探讨人类认知的复杂性?
人类认知的本质是一个复杂的问题,在不同的领域有不同的答案,并且引起矛盾和零散的理论,就像古印度耆那教寓言中的大象阐明了个人知觉的相对现实,如同耆那教非绝对论哲学系统所形容1。一头大象抵达一个盲人村落,当村民试著要猜想是什么莅临村落时,他们提出了一个截然不同的假设。摸到象脚的盲人认为它是一棵树,而抓住大象尾巴的盲人认为它是一根绳子。没有一个盲人能够独自弄清楚大象是什么,只有汇集大家不同的经验才有办法认识大象。人类的认知就是认知科学中的大象,而盲人就是各个学科——哲学、心理学、神经科学、语言学、计算机科学、人类学和文学理论——都曾经徒劳无功的分别尝试理解我们的心理如何运作。
20世纪初,现代控制论的出现反映了人们渴望透过结合跨领域的知识,同时将“控制”的概念置于中心位置,以建立一种新的沟通模式的研究领域。直到四〇年代在纽约举行的梅西会议(Macy Lectures)2,将数学家、哲学家、人类学家、心理学家和经济学家聚集在一起,认知的研究才在国际上有了新的方向。这些会议以及许多关于心智本质的理论辩论,引发了新的势力,和一个新兴学科——计算机科学的发展相呼应,计算机科学的潜在应用前景促成了“计算机脑”的隐喻热潮。
然而,这种被称为“计算主义”的隐喻并没有被整个科学界接受。除了演算法的效率,控制论者及其后继者坚持自组织过程在认知过程中出现的重要性。不过,计算机模型并没有考虑到大脑功能的所有属性。实际上,这些属性最初是为了计算机概念而预留的。讽刺的是,计算机之父艾伦·图灵(Alan Turing )和约翰‧冯‧诺伊曼(John Von Neumann)的最新著作透过大脑功能的自组织方面,强调了这一问题,而大脑在计算机中是完全没有的。
在第二次控制论浪潮的思想家中,智利哲学家、生物学家温贝托‧马图拉纳(Humberto Maturana)和神经科学家弗朗西斯科‧瓦雷拉(Francisco Varela)与认知主义的工作相辅相成,对某些现象,特别是大脑重构现实世界的过程进行深入的研究,例如记住展览中看到的美、看画的顺序等等。这二位智利专家透过强大的生物学知识来看待这些现象。在更具体的思考认知之前,我们试图从一个新的角度来概念化人生。1970年代,分子生物学蓬勃发展,细胞越来越多地像一个复杂的机器出现在生物学家面前,这二位神经生物学家把思考的重点放在生命的二个中心和互补方面——自主性和与环境的结合。
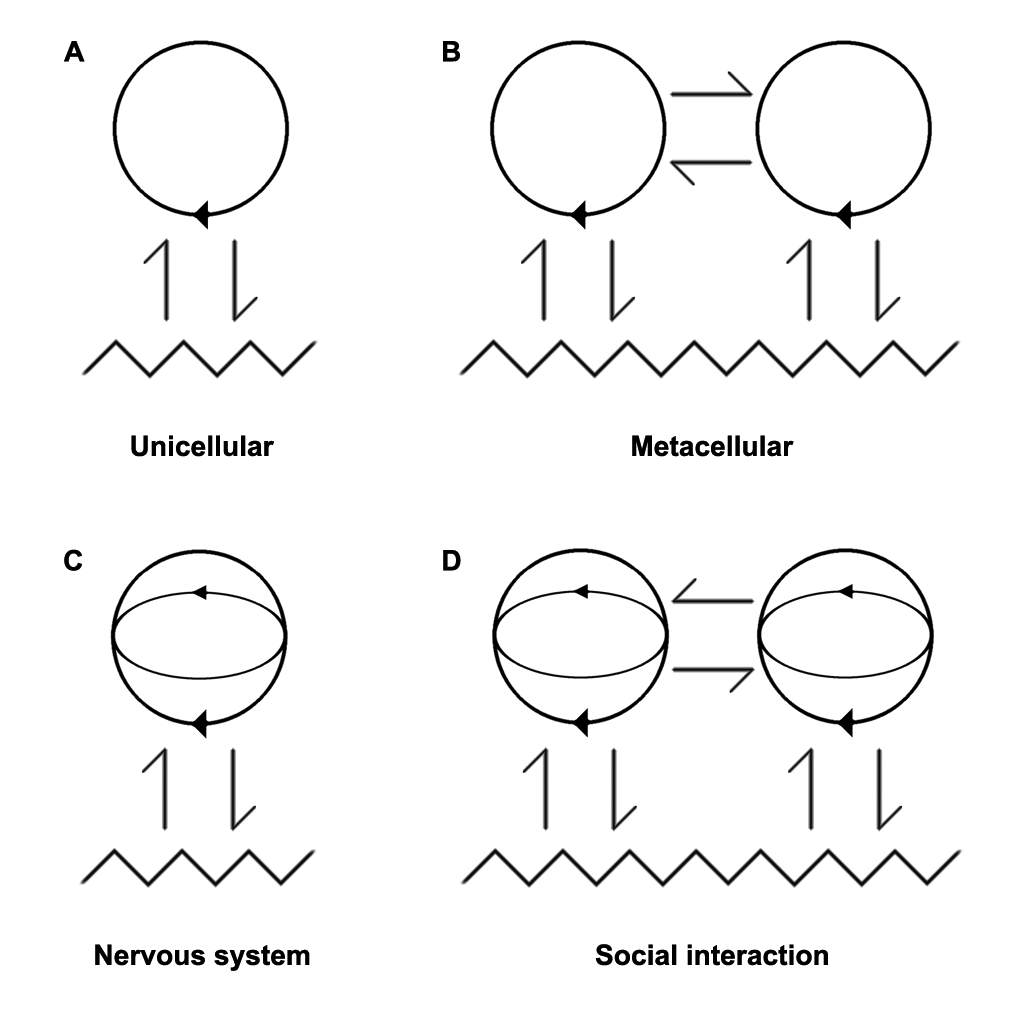
马图拉纳和瓦雷拉提出了自生系统论(autopoiesis)-auto在希腊语中意为“自我”而poiesis意为 “产生”(图拉纳和瓦雷拉,1994)细胞被认为是生命的基本单位,在这个基本的组织层次上,已经具备了建立自主结构的必要条件,但又与环境相结合。这种结构上的耦合——被称为“一阶”耦合——是一个封闭在因果网路而维持细胞自我的组织或复制。
这个理论框架有助于设想在进化过程中,稳定与环境的结构耦合如何为建立细胞本身之间的耦合提供必要条件(图一)。这种从单细胞到多细胞实体的转变应于5.41亿年前的寒武纪爆发3 。在地球生命史上,这一里程碑见证了最早的真核生物(其细胞具有细胞核的单细胞生物和多细胞生物)、原生动物(protozoa,proto在希腊语中是第一的意思,zone是动物)变成后生动物,其细胞可以发挥不同的功能从而造福整个生物体系。这种专业化使得具有组织水准又复杂许多的生物系统出现,从而使营养自主性以外的过程得以发展。因此,我们看到免疫系统的出现,它确保生物体对疾病的防御,但最重要的是神经系统,它使我们能够感知环境并对环境做出更好的反应。
结构耦合就成了“二阶”,因为这种与环境的循环不再只是维持生物体的物质自主性,还保持了信息自主性。瓦雷拉后来发展出生成论,特别关注这些方面4,不再关注生物的进化和结构,而是关注认知。如同他提出的自生系统论一样,瓦雷拉拒绝使用计算机大脑的隐喻,并玩弄自主性和耦合的想法,从而与同时代的人拉开了距离。
由于细胞和环境耦合演化过程中的稳定化为细胞自身的耦合提供了必要的条件,因此生物体与其环境之间信息互动的稳定化使这些生物之间能够相互作用(图一)。这种“三阶”结构耦合无非是社会互动,是文化出现的先决条件。
理解社会互动:神经科学的新挑战
尽管看起来很矛盾,但社会神经科学的工作长期以来一直集中于孤立的大脑上,导致一些研究人员使用另一个比喻,将社会互动视为社会神经科学的“暗物质 ”5 (Schilbach等,2013)。直到2000年代,神经科学家才认真对待社会互动,研究的不仅是一个大脑,而是两个大脑的共力作用(Hari & Kujala,2009)。
因此必须面对二个挑战。在道德层面上,必须发明实验性协议来质疑这种互动的尺度,在技术层面上,必须建立工具来即时记录双向交流的大脑活动。在此之前,对大脑的研究一直以社会知觉为导向,建立专注于研究社会刺激的感知,这些研究并未考虑到交流的对等性和共同协调性。例如,参与者必须从面部图像中检测出一种情绪,或模彷影片中成和的场景。但很显然的是,由于面部图像或影片对观众的行为仍不敏感,这种情况下的讯息是单向,而不是社会互动中的双向讯息。
我们如何才能关闭自我和他人互相影响的循环?三种主要方法已经被研发出来:一、同时研究几个人类“活体”。二、用数学模型做“电脑模拟” 的相互作用。三、结合这二种方法,观察人和数学模型之间的互动。
相同波长的大脑,或探索同步
在心理学中已经研究了几个人类参与者的相互作用,例如在儿童发展研究中,强调婴儿与其父母之间交流的基本作用(Fogel, 1993)。然而,要将这种方法应用于神经科学,必须与参与互动者的大脑活动纪录相结合。
这就是被称为“超扫描”方法的目的,这个方法于2002年引入磁共振功能成像(fMRI; Montague等,2002),然后在2006年扩展到脑电图(EEG)(Babiloni等,2006)6 。这些最初的研究证明了同时记录的可行性,并在最初的结果证实了我们的大脑在共享相同讯息时,往往会在社交环境中做出类似的反应。然而,第一批超扫描记录无法显示三阶结构耦合——即根据一个共同的外部刺激(例如,他们跳舞的音乐或他们同时看的电影),互动参与者的大脑会同步。
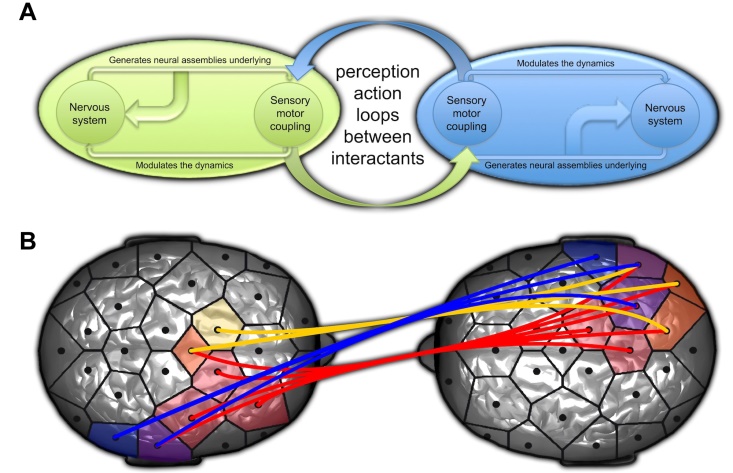
透过将自发性模彷(一种发展心理学任务,二个人自由模彷对方)和脑电图记录结合,我们已经能够实现这种三阶耦合。当参与者自发的互动时,他们会进入同步状态,即他们的行为在时间上是彼此依赖的,以至于他们的动作同时开始和结束,即使这些动作不完全相同。透过独立这些自发性模彷手部动作过程中的互动同步时刻,我们已经能证明它们伴随著参与者大脑之间的脑电波同步(Dumas等,2010;图二)。
这个结果除了与术语“同相”或“处于同一波长”一致,还表明人类大脑对即时社交互动的反应方式与社交感知的反应不同。例如,在自发性的双向互动(例如对话)过程中,大脑奖励迴路的活化更多,当他们被动地观看著独白时,大脑奖励迴路的活化较少。因此强调这些变化,就对以往社会神经科学领域的结果,仅基于对社会知觉的单向研究提出质疑。结果还强调了脉络与社会互动中所扮演的角色(例如模彷者或原型)在调节大脑反应中的重要性(Nadel&Dumas,2014)。这些新的方法可能会应用于如何设计更多互动学习和表演艺术。
社会互动的双面性
因此,我们看到了了解人类认知的方法可以被更新。超越计算机大脑的隐喻,人类的思想可以被看作是体内生物嵌入(尤其是大脑)和社会动态(Dumas, 2011)之间的一个介面。在二个介面之间存在著双重限制:社交大脑使我们能够与其他大脑进行讯息耦合,而社会互动塑造了我们的大脑连结(Clark and Dumas,2016)。因此就像盲人和大象的神话一样,认知科学的不同学科似乎必须跨领域和观察层次以共同建构意义(Dumas, Laroche,Lehmann,2014)。社会互动似乎是我们个体间的动力和个体生物基础间的精巧混合。
译自本刊英文版原文《On The Complementarity Of Inter-individual Social Dynamics And Intra-individual Biological Grounding》
- 起源于印度的哲学体系,在我们这个时代之前就发展起来,认为所有现实都是相对的,并存在多种观点。
- 1942年到1953年间,在神经学家沃伦·麦卡洛克(Warren McCulloch)的倡议下,在纽约举办了两次会议。这些会议是接受梅西基金会的财务支持而命名的。
- 数千万年的时期(也被称为生态大爆炸),在此期间可以观察到动物、植物和细菌种类的极大多样化。
- 生成(Enaction)是生物与环境互动时自身组织的方式。
- 隐喻指一种物理物质,也称为暗物质,它是由实验室中未探测到的粒子组成,但根据某些天体物理学观察,这些粒子代表宇宙中相当大的质量。
- 关于这个轶事,在六〇年代就用脑电图EEG进行实验,但是在超心理学领域。
参考书目
Maturana, Humberto R., Francisco J. Varela, & François-Charles Jullien. L’arbre de la connaissance. Ed. Addison-Wesley France, 1994.
Schilbach, L., Timmermans, B., Reddy, V., Costall, A., Bente, G., Schlicht, T., & Vogeley, K. (2013). Toward a second-person neuroscience 1. Behavioral and brain sciences, 36(4), 393-414.
Hari, R., & Kujala, M.V. (2009). Brain basis of human social interaction: from concepts to brain imaging. Physiological reviews, 89(2), 453-479.
Montague, P.R., Berns, G.S., Cohen, J.D., McClure, S.M., Pagnoni, G., Dhamala, M., Wiest, M.C., Karpov, I., King, R.D., Apple, N., & Fisher, R.E. (2002). Hyperscanning: simultaneous fMRI during linked social interactions. Neuroimage, 16(4), 1159- 1164.
Babiloni, F., Cincotti, F., Mattia, D., Mattiocco, M., Fallani, F. D. V., Tocci, A., Bianchi, L., Marciani, M.G., & Astolfi, L. (2006). Hypermethods for EEG hyperscanning. In Engineering in Medicine and Biology Society, 2006. EMBS’06. 28th Annual International Conference of the IEEE, 3666-3669.
Dumas, G., Nadel, J., Soussignan, R., Martinerie, J., & Garnero, L. (2010). Inter- brain synchronization during social interaction. PloS one, 5(8), e12166.
Nadel, J., & Dumas, G. (2014). The interacting body: intra-and interindividual processes during imitation. Cogn. Educ. Psychol, 13, 163-175.
Dumas, G. (2011). Towards a two-body neuroscience. Communicative & integrative biology, 4(3), 349-352.
Clark, I., & Dumas, G. (2016). The regulation of task performance: a trans- disciplinary review. Frontiers in psychology, 6, 1862.
Dumas, G., Laroche, J., & Lehmann, A. (2014). Your body, my body, our coupling moves our bodies. Frontiers in human neuroscience, 8, 1004.
法国巴斯德中心人类遗传学与认知功能实验室研究员,美国佛罗里达大西洋大学(Florida Atlantic University)复杂系统与脑科学中心 “人类大脑与行为实验室 ”成员。吉拥‧都马还是巴黎罗伯特-德布雷医院(hôpital Robert-Debré)青少年与儿童精神病学科SoNeTAA平台的创始人与科学协调员。
法国巴斯德中心人类遗传学与认知功能实验室研究员,美国佛罗里达大西洋大学(Florida Atlantic University)复杂系统与脑科学中心 “人类大脑与行为实验室 ”成员。吉拥‧都马还是巴黎罗伯特-德布雷医院(hôpital Robert-Debré)青少年与儿童精神病学科SoNeTAA平台的创始人与科学协调员。
